Оглавление
- 1 Что такое филогенетический ряд
- 2 Параллелизмы и их эволюционное значение
- 3 Доказательства эволюции живой природы. Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
- 4 Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
- 5 Немного о ДНК
- 6 Основные принципы построения филогений
- 7 Биогеографические доказательства эволюции
- 8 Сходство зародышевого развития позвоночных
- 9 Типы деревьев
- 10 polytomies
- 11 Дикие лошади
- 12 Доказательства эволюции живой природы
Деревья состоят из узлов, которые соединяются с «ветвями». Конечные узлы каждой ветви представляют собой терминальные таксоны и представляют последовательности или организмы, для которых известны данные — это могут быть живые или вымершие виды.
Что такое филогенетический ряд
Вопрос 1. В чем различие макро- и микроэво-люции?
Под микроэволюцией мы понимаем образование новых видов.
Понятием макроэволюции обознача-ют происхождение надвидовых таксонов (рода, отряда, клана, типа).
Тем не менее нет принципиальных раз-личий между процессами образования но-вых видов и процессами формирования более высоких таксономических групп. Термин «микроэволюция» в современном смысле был введен Н. В. Тимофеевым-Ресовским в 1938 г.
Вопрос 2. Какие процессы являются движу-щими силами макроэволюции? Приведите приме-ры макроэволюционных изменений.
В макроэволюции действуют те же про-цессы, что и при видообразовании: об-разование фенотипических изменений, борьба за существование, естественный отбор, вымирание наименее приспособ-ленных форм.
Результатом макроэволюционных про-цессов становятся существенные измене-ния внешнего строения и физиологии орга-низмов — такие, например, как формиро-вание замкнутой системы кровообращения у животных или появление устьиц и эпи-телиальных клеток у растений. К фун-даментальным эволюционным приобрете-ниям такого рода относятся образование соцветий или превращение передних ко-нечностей рептилий в крылья и ряд дру-гих.
Вопрос 3. Какие факты лежат в основе изуче-ния и доказательств макроэволюции?
Наиболее убедительные доказательства макроэволюционных процессов дают нам палеонтологические данные. Пале-онтология изучает ископаемые остатки вымерших организмов и устанавливает их сходство и различия с современными организмами. По остаткам палеонтологи реконструируют внешний облик вымер-ших организмов, узнают о растительном и животном мире прошлого. К сожале-нию, изучение ископаемых форм дает нам неполную картину эволюции флоры и фауны. Большинство остатков состоит из твердых частей организмов: костей, раковин, внешних опорных тканей расте-ний. Большой интерес вызывают окаме-нелости, сохранившие на себе следы нор и ходов древних животных, отпечатки конечностей или целых организмов, ос-тавленных на когда-то мягких отложе-ниях.
Вопрос 4. Какое значение имеет исследова-ние филогенетических рядов? Материал с сайта
Исследование филогенетических ря-дов, построенных на основе данных па-леонтологии, сравнительной анатомии и эмбриологии, важно для дальнейшего развития общей теории эволюции, постро-ения естественной системы организмов, воссоздания картины эволюции конкрет-ной систематической группы организмов.
В настоящее время для построения фи-логенетических рядов ученые все больше привлекают данные таких наук, как гене-тика, биохимия, молекулярная биоло-гия, биогеография, этология и др.
Эти физические перемены произошли одновременно с серьёзными изменениями в популяционной плотности и социальной структуре. Специалисты называют их «эволюционным новшеством, новым свойством, отсутствовавшим у предковой популяции и развившемся в ходе эволюции у этих ящериц».
Филогене́з (от греч. «phylon» — род, племя и «генез»), историческое развитие организмов, в отличие от онтогенеза- индивидуального развития организмов. Филогенез — эволюцию в прошлом — невозможно наблюдать непосредственно, а филогенетические реконструкции невозможно проверить при помощи эксперимента.
Например, еще в 1844 году были найдены какие-то окаменевшие зубчики, названные конодонтами. На челюстях, в отличие от всех современных птиц, были зубы, как у пресмыкающихся. Второе затруднение состоит в том, что полностью изучить организацию даже одноклеточного организма технически невозможно. И у самых примитивных четвероногих, и у двоякодышащих рыб есть легкие и трехкамерное сердце, состоящее из двух предсердий и одного желудочка.
Параллелизмы и их эволюционное значение
Эта ветвь эволюции низших позвоночных возникла в конце кембрийского периода и в ископаемом состоянии неизвестна с конца девонского периода. Дело в том, что у ископаемых бесчелюстных обызвествлялись жаберные полости, полость головного мозга, стенки многих крупных кровеносных сосудов и других внутренних органов. В этом случае работает только сравнительная анатомия и в очень необходимой степени — эмбриология. Широкое распространение компьютерной техники облегчило такой анализ, и кладограммы (от греч. «klados» — ветвь) стали появляться в большинстве филогенетических публикаций.
Исследование структуры нуклеиновых кислот и других макромолекул стало в настоящее время одним из важнейших дополнений к методу тройного параллелизма. Это можно было бы принять за ошибку, если бы в 1983 году М. Ф. Ивахненко не доказал на палеонтологическом материале, что черепахи произошли от амфибий независимо от всех остальных пресмыкающихся.
Уточнение выражается в том, что реконструкции становятся все более подробными. Если в окружающем нас мире есть что-то неизвестное, то задача науки — изучить и объяснить это неизвестное, независимо от теоретического и практического значения предмета изучения. Кроме того, филогенетические реконструкции являются основой, на которой выясняются закономерности эволюции.
Существует еще много и других закономерностей эволюции, которые были обнаружены благодаря филогенетическим исследованиям. Эволюционные процессы наблюдаются как в естественных, так и лабораторных условиях. Факт эволюции на внутривидовом уровне доказан экспериментально, а процессы видообразования непосредственно наблюдались в природе.
Кроме мустангов в Северной Америке есть два вида диких островных пони — на островах Ассатиг и Сейбл. Полудикие табуны лошадей камаргу встречаются на юге Франции. В горах и болотах Британии тоже можно встретить некоторых диких пони.
Доказательства эволюции живой природы. Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
Биологами, палеонтологами и представителями других отраслей естествознания получены многочисленные свидетельства, иллюстрирующие и доказывающие эволюционную теорию Дарвина. Наиболее показательны палеонтологические, сравнительно-анатомические, сравнительно-эмбриологические и биогеографические доказательства эволюции.
Эволюционная теория со времени Ч. Дарвина обогатилась данными палеонтологии, анатомии, биогеографии и других наук.
Палеонтологические доказательства
Ископаемые вымершие организмы находили и до создания теории эволюции. Собраны окаменевшие отпечатки, пропитанные кремнеземом, известняком остатки, следы жизнедеятельности. К редким находкам относятся хорошо сохранившиеся организмы, вмерзшие в лед, «законсервированные» в вечной мерзлоте, заключенные в янтарь.
Найдены промежуточные формы (переходные), сочетающие признаки эволюционно древних и более молодых групп организмов. К таковым относятся отпечатки растений псилофитов, окаменевшие остатки стегоцефалов, археоптерикса, зверозубых ящеров (Таблицы 1, 2 и 3, рис. 2), «живое ископаемое» — рыба латимерия (рис. 1).
Псилофиты — переходная форма между водорослями и наземными растениями
Признаки наземных растений
- Тело — слоевище, не дифференцированное на органы;
- Ткани отсутствуют.
- Стебель с чешуйками;
- Устьица в кожице стебля и листьев;
- Корневище с ризоидами;
- Есть покровные, механические и проводящие ткани.
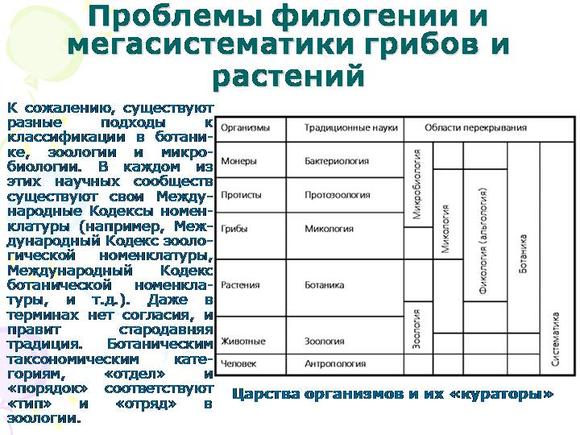
Рис. 1 Кистеперая рыба латимерия
Археоптерикс
Археоптерикс — промежуточная форма между пресмыкающимися и птицами
- Длинный хвост с позвонками;
- Пальцы с когтями;
- Наличие зубов;
- Отсутствие киля;
- Брюшные ребра.
- Внешнее сходство;
- Наличие крыльев;
- Оперение;
- Сходство задних конечностей с цевкой.
Рис. 2 Археоптерикс
Зверозубый ящер
Зверозубый ящер — переходная форма между пресмыкающимися и млекопитающими
- Внешний облик;
- Расположение конечностей по бокам туловища, а не под ним.
- Строение черепа, позвоночника и конечностей;
- Деление зубного ряда на клыки, резцы и коренные зубы.
Значительным доказательством эволюции считается то, что в более ранних слоях встречаются просто устроенные организмы, по сравнению с находками в более поздних субстратах. Останки видов, обитавших в одни геологические эпохи, позже исчезают и больше не появляются. Это означает, что виды возникают в ходе эволюции и вымирают в последующие эры и периоды.
Сравнительно-анатомические доказательства
Находки ископаемых останков организмов часто имеют общее происхождение и сходство с современными. Изучение строение ныне существующих и исчезнувших позвоночных животных выявило общие черты строения:
- Двусторонняя симметрия.
- Твердый скелет.
- Полости тела.
- Сходное строение нервной системы.
Результаты эволюции: приспособленность организмов к среде обитания, многообразие видов
Виды живых организмов характеризуются общими признаками и удивительным разнообразием внешнего и внутреннего строения, процессов жизнедеятельности, поведения. Все черты появились в процессе эволюции, подверглись действию естественного отбора. В конечном итоге разные изменения обеспечили лучшее приспособление организмов к среде обитания.
Различают физиологические приспособления и особенности размножения, обусловленные адаптацией. Есть разнообразные морфологические черты приспособленности к условиям обитания и особенности поведения, облегчающие выживание.
- покровительственная окраска;
- предостерегающая окраска;
- мимикрия (рис. 7);
- маскировка.

Рис. 7. Мимикрия и покровительственная окраска у насекомых
Мимикрия — способность организмов принимать форму, подражать окраске и особенностям поведения других организмов и неживых тел. Например, беззащитные животные подражают хищным. Такие особенности дают преимущества в борьбе за выживание.
Покровительственная окраска позволяет слиться с окружающей средой. Хищникам такая особенность нужна для более успешной охоты, жертвам — для того, чтобы не быть съеденными.
Предостерегающая окраска характерна для ядовитых организмов. Яркие цвета и оттенки предупреждают потенциальных хищников о несъедобности жертвы. Перечисленные приспособления закреплены естественным отбором, как обеспечивающие выживание вида в борьбе за существование.
Колючки, шипы, жгучие волоски, гликозиды в тканях — приспособления растений, защищающие их от поедания животными. Эпифитные растения могут обойтись без почвы. Они поглощают влагу из воздуха при помощи воздушных корней. Насекомоядные растения получают азот на бедных почвах, способны переваривать насекомых.
Увеличение шансов на выживание дают многие приспособления у живых организмов. Появление адаптаций у видов происходило в течение больших промежутков времени в процессе борьбы за существование. Однако приобретенные черты полезны и целесообразны в конкретной среде обитания; они могут оказаться ненужными, вредными в других условиях.
Состав живых организмов описывают следующие показатели. Всего на Земле насчитывается свыше 2 млн видов. Около 1,5 млн видов животных, порядка 400 тыс. видов растений, 100 тыс. видов грибов и бактерий. Поразительное разнообразие — результат дивергенции. Виды расходятся по морфофизиологическим, биохимическим, репродуктивным, генетическим и экологическим признакам.
Классификация организмов — задача систематики. Со времени работ основателя науки К. Линнея, ученые стараются отразить степень родства организмов. Тогда классификация будет носить максимально естественный характер.
Самыми основными деревьями являются кладограммы. Эти графики показывают отношения с точки зрения общих предков (согласно самым последним общим предкам).
Немного о ДНК
Чтобы понять, как анализируют ДНК, надо вспомнить, как она устроена. ДНК, или дезоксирибонуклиновая кислота, — это очень длинная молекула, которая находится в ядре клетки. ДНК, как правило, состоит из двух закрученных спиралей, а каждая спираль состоит из множества нукеотидов. Нуклеотиды по большей части отличаются друг от друга азотистыми основаниями, которых в ДНК всего четыре: аденин, тимин, гуанин и цитозин. Именно нуклеотиды создают слабые химические связи, которыми соединяются спирали ДНК. Аденин одной спирали связывается с тимином другой спирали, а гуанин связывается с цитозином (рис. 1). Мутация происходит, когда одно основание заменяется на любое другое. Чаще всего замены происходят в парах аденин—гуанин и тимин—цитозин.
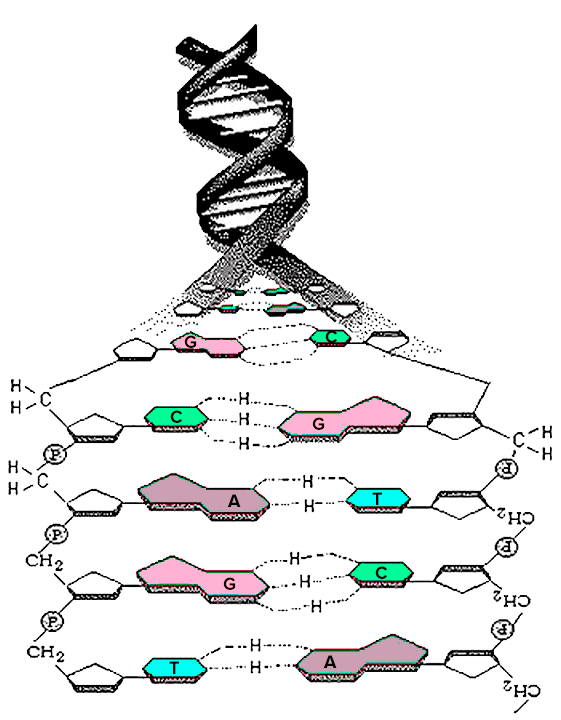
Рисунок 1. ДНК. A — аденин, C — цитозин, G — гуанин, T — тимин.
В ДНК есть последовательности нуклеотидов, которые кодируют белки, и есть участки, которые ничего не кодируют. Кодирующие последовательности — это гены. Они могут быть разной длины, но чаще всего имеют определенную структуру, по которой можно сказать — ген это или нет. Именно гены обычно используют для филогенетического анализа.
Основные принципы построения филогений
Наверное, самое главное правило, которым руководствуются для построения филогений в наше время — это принцип дихотомии: считается, что из трех таксонов, два должны быть более родственны друг другу, чем третий. Поэтому филогении обычно выглядят как дихотомически разветвленные деревья. Если порядок ветвления установлен для всего дерева, то говорят, что оно полностью разрешенное. Иногда в филогениях бывают «кусты» или политомии — это те места, где порядок ветвления неясен, тогда говорят, что дерево не полностью разрешенное. Этот принцип несовершенен, потому что эволюция таксонов далеко не всегда происходит дихотомически. Когда становится понятно, что дихотомия не отражает реальный случай, исследователи привлекают другие схемы — например, филогенетические сети 8.
Методы построения филогений еще в 60-х годах XX века разделились на две основные ветви — фенетические и кладистические. В то время анализ родственных связей основывался на морфологических признаках 12; с привлечением к построению филогений молекулярных признаков основные принципы анализа родственных связей остались фактически теми же.
- В фенетике построение филогении основано на общем сходстве двух видов — то есть, чем больше общих признаков, тем ближе они друг к другу;
- В кладистике же считается, что только уникальные для какой-либо группы признаки можно использовать для оценки родства таксонов. Родоначальником кладистического анализа является немецкий ученый Вилли Хенниг 6. Этот автор также ввел и терминологию, которая широко используется до сих пор. Уникальные признаки называются апоморфиями; ветви, которые объединяются апоморфиями — это клады; а сама филогения называется кладограммой (рис. 2) 12.
Чтобы было более понятно, представьте три вида животных: домашнюю мышь, сумчатую мышь и кенгуру. Домашняя мышь и сумчатая мышь очень похожи друг на друга внешне, но у сумчатой мыши и кенгуру есть общая апоморфия — сумка, — что говорит о том, что эти два вида родственные. Но, естественно, филогенетический анализ основывается на гораздо большем количестве признаков, и группы могут иметь несколько апоморфий.
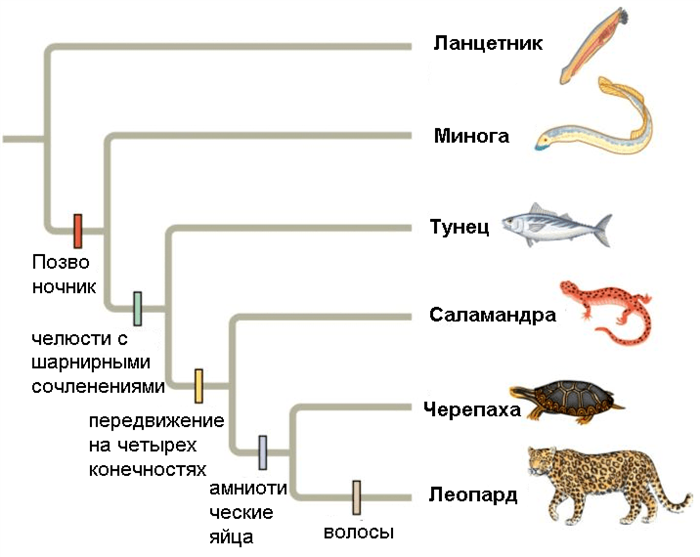
Рисунок 2. Полностью разрешенная кладограмма. Каждое ветвление — это клада. Обозначенные признаки являются апоморфиями.
Есть два типа политомий. Первыми являются «жесткие» политомии. Они присущи исследовательской группе и указывают на то, что потомки развивались одновременно. Альтернативно, «мягкие» политомии указывают на неразрешенные отношения, вызванные данными per se.
Дивергенция
Дивергенция начинается с внутривидовой дифференциации популяций. Обычно она приводит к формированию более чем двух подвидов. Рассматривая видообразование, специалисты объясняют расхождение близких видов по разным нишам межвидовой конкуренцией в сочетании с изоляцией. Однако согласно экологическим исследованиям межвидовая конкуренция проявляется прежде всего при инвазиях — вселениях видов, новых для данной экосистемы. При этом в сложившихся сообществах часто не удается выявить межвидовую конкуренцию даже у экологически близких видов. Возможно, это объясняется несовершенством методов полевой экологии. При экспериментальном изъятии одного из потенциальных видов-конкурентов или сокращении его численности другой вид занимает его нишу. Так произошло, например, в Йеллоустонском национальном парке, где из лучших побуждений уничтожили волков. Это привело к повышению смертности оленей от разных болезней и к тому, что волков заместили размножившиеся койоты. Пришлось реакклиматизировать волков, чтобы восстановить баланс экосистемы.
Если говорить о макроэволюции, то конкуренции между надвидовыми таксонами придают очень большое значение. Выше уже упоминалась роль конкуренции в формировании границ адаптивных зон. Так же как внутри видов может формироваться несколько подвидов, при освоении новой адаптивной зоны начинается адаптивная радиация 1 : формируется сразу несколько направлений эволюции, которые приводят к возникновению нескольких дочерних таксонов более низкого ранга. Среди сухопутных хищных млекопитающих в биосфере Земли насчитывается 12 семейств, с моржами и тюленями — 15. Таксон, осваивающий новую адаптивную зону, освобождается от конкуренции с таксонами, оставшимися в адаптивной зоне предков, и не испытывает конкуренции с аборигенами новой зоны (или побеждает в ней). В результате идет дифференциация дочерних таксонов, которая продолжается до тех пор, пока они не заполнят новую зону и не поделят ее на субзоны. Деление адаптивной зоны на субзоны означает переход от дивергенции к параллельной эволюции дочерних таксонов.
Возможен и другой вариант. Какие-то ветви адаптивной радиации могут приспосабливаться к условиям существования, нетипичным для занимаемой адаптивной зоны. К этому их могут подталкивать и конкуренция, и наличие недоиспользуемых ресурсов. Так, моржи и ушастые тюлени произошли из того же корня, что и современные медведи. Адаптивная зона (уже не субзона) этих двух семейств формировалась иначе, чем субзона сухопутных хищников. Дивергенция продолжалась до тех пор, пока не остановилась на границе зоны настоящих тюленей. Можно предполагать, что теперь эволюция морских котиков и морских львов пойдет по пути дальнейшей адаптации к жизни в море. Мех котиков служит надежной теплоизоляцией, но мешает глубоко нырять: прослойка воздуха, удерживаемая мехом, создает дополнительную плавучесть. Если нырять глубоко, давление воды выдавливает воздух из меха и он промокает, то есть теряет функцию термоизоляции. У настоящих тюленей шерсть короткая и жесткая. Она годится только для механической защиты кожи. Термоизоляцию обеспечивает подкожный жир. Словом, можно предположить, что дивергенция рано или поздно обязательно сменяется параллелизмом.
Параллельная эволюция
После дробления адаптивной зоны на субзоны, когда дивергенция прекращается, каждый таксон продолжает адаптироваться к условиям своей новой субзоны. Наступает период параллельной эволюции. На общей основе, унаследованной от предков, начинают формироваться сходные адаптации. Это обусловлено тем, что субзоны — части общей адаптивной зоны и условия существования в них во многом одинаковы. Ярким примером параллелизма могут служить неправильные морские ежи.
Исходно морские ежи, как и морские звезды, — хищники. Их скелет состоит из известковых пластинок пятилучевой симметрии, совместно образующих полусферу выпуклостью вверх. Рот расположен на нижней плоской стороне полусферы, а анальное отверстие — в центре выпуклой верхней стороны. Неправильные морские ежи в юрском периоде приспосабливались к питанию илом. Главное преимущество ила в том, что его много. В нем достаточно органики, пригодной для питания, но хватает и грязи. Питание илом потребовало поглощения больших количеств этого субстрата, а тем самым удлинения пищеварительного тракта. В пяти группах морских ежей это привело к смещению анального отверстия назад. В результате тело (скелет) изменило форму, стало вытянутым. Сформировались неправильные морские ежи (рис. 54).
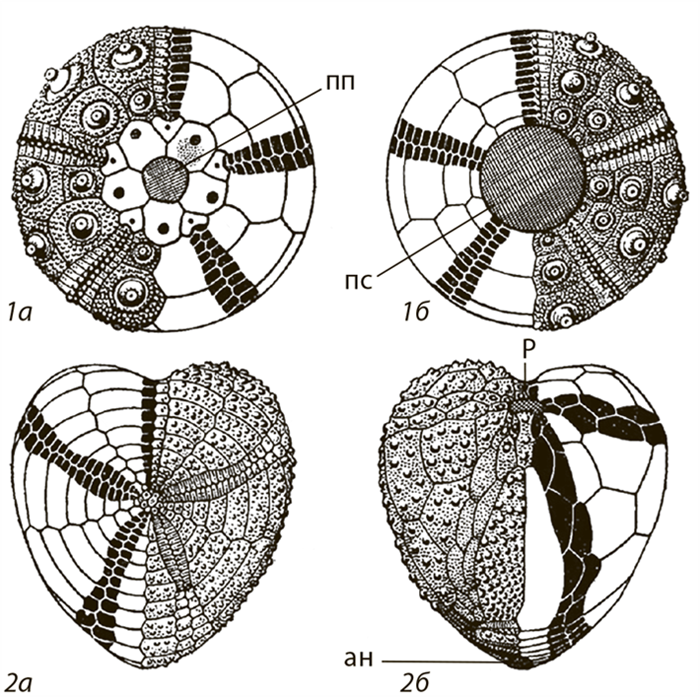
Рис. 54. Схема строения панциря правильных (1) и неправильных (2) морских ежей (из Друщица, 1974) (а — вид сверху, б — вид снизу): ан — анальное отверстие; пп — перипрокт (небольшое поле, посередине которого расположено заднепроходное отверстие); пс — перистом с ротовым отверстием; р — рот
Чем больше сходство адаптивных зон или субзон, тем глубже сходство параллельно эволюционирующих признаков. Например, двояковогнутые позвонки — их можно увидеть у рыбы на вашем столе — возникли у двоякодышащих рыб, у костных ганоидов (малоизвестная широкой публике группа лучеперых рыб), у костистых рыб и у амфибий. Наиболее глубокий параллелизм наблюдается в эволюции тканевого уровня организации. Поперечнополосатая мускулатура есть у позвоночных и у насекомых. В более примитивном состоянии, без оформленных мышечных волокон, она есть даже у медуз. Крупный отечественный гистолог А. А. Заварзин выявил глубочайший параллелизм в эволюции нервной ткани при сравнении первично- и вторичноротых животных: многощетинковых червей и личинок миног, насекомых и позвоночных.
Параллелизмы затрудняют филогенетические реконструкции и часто служат причинами ошибок в них. Неполнота палеонтологической летописи приводит к тому, что в один филогенетический ряд попадают организмы из разных параллельных таксонов. Сходство между ними высокое, а родство отдаленное.
Замечу, что параллелизмы не всегда развиваются одновременно. В нескольких случаях доказано, что в одной из групп они возникли раньше, чем в другой. Тогда пишут о «филогенетическом предварении», «пророческих группах» и т. п. Эти факты охотно используют противники эволюции на основе естественного отбора. Ведь если два таксона приобретают одинаковые признаки, причем один — раньше, а другой — позже, то отбор тут ни при чем. Мол, действуют иные законы эволюции, направляющие ее по заранее намеченному пути.
Примером асинхронного (не одновременного) параллелизма может служить эволюция южноамериканских непарнокопытных Litopterna и лошадей, включая зебр и ослов. Литоптерны были однопалыми, как и лошади. Эволюция обоих отрядов шла одинаково. Редуцировались боковые пальцы, пока не остался только средний с копытом. Литоптерны существовали с позднего палеоцена и вымерли в плейстоцене. Эволюция современных лошадей началась в эоцене.
Различают физиологические приспособления и особенности размножения, обусловленные адаптацией. Есть разнообразные морфологические черты приспособленности к условиям обитания и особенности поведения, облегчающие выживание.
Биогеографические доказательства эволюции
Сравнение фаун и флор разных континентов. Яркое свидетельство эволюционного процесса — распространение животных и растений по поверхности нашей планеты. Во все времена путешественников и натуралистов поражало своеобразие растительного и животного мира Австралии, Южной Америки, океанических островов и вызывало интерес сходство фауны некоторых материков, например Северной Америки и Евразии. А. Уоллес привел все сведения в систему и выделил шесть зоогеографических областей:
1) Палеоарктическую, охватывающую Европу, Северную Африку, Северную и Среднюю Азию, Японию;
2) Неоарктическую, включающую Северную Америку;
3) Эфиопскую, включающую Африку к югу от пустыни Сахара;
4) Индомалайскую, охватывающую Южную Азию и Малайский архипелаг;
5) Неотропическую, занимающую Южную и Центральную Америку;
6) Австралийскую, включающую Австралию, Новую Гвинею, Новую Зеландию, Тасманию, Соломоновы острова и Новую Каледонию.
Степень сходства и различия между разными зоогео-графическимн областями неодинакова. Фауна и флора Палеоарк-тической и Неоарктической областей имеют много общего, хотя и изолированы Беринговым проливом- Неоарктическая и Неотропическая области, наоборот, существенно отличаются, хотя и соединены сухопутной связью (Панамским перешейком). В чем же причины этого сходства и различия? Очевидно, они связаны с историей формирования материков, временем их изоляции. Так, глубокое различие в фауне Неотропической и Неоарктической областей определяется тем, что сухопутная связь между ними установилась совсем недавно. Об этом свидетельствуют геологические данные.
После возникновения Панамского моста лишь немногим южноамериканским видам удалось проникнуть на север (например, дикобраз, броненосец, опоссум). Североамериканские виды преуспели в освоении южноамериканской области несколько больше. Олени, лисы, выдры, медведи проникли в Южную Америку, но не оказали существенного влияния на ее уникальный видовой состав. Только здесь живут представители отряда неполнозубых (муравьеды и ленивцы), сохранились птицы гоацины, которые могут лазать по деревьям благодаря когтям на пальце крыла (как археоптерикс), и другие животные. Своеобразие фауны Южной Америки было одним из фактов, натолкнувших Ч. Дарвина на мысль об эволюции. Сходство фауны Неоарктической и Палеоарктической областей обусловлено тем, что в прошлом между ними существовал сухопутный мост — Берингов перешеек.
Наиболее отличен от других континентов животный мир Австралии. Известно, что Австралия обособилась от Южной Азии свыше 100 млн. лет назад, еще до возникновения высших млекопитающих. Лишь в Ледниковый период сюда через острова Зонд-ского архипелага перебрались немногие плацентарные — мыши и собаки (потомок последних—австралийская собака динго). В остальном фауна Австралии очень своеобразна.
Таким образом, чем теснее связь континентов, тем более родственные формы там обитают, чем древнее изоляция частей света друг от друга, тем больше различия между их населением.
Видовой состав фауны и флоры островов целиком определяется историей их происхождения. Острова могут быть материковыми, представлять собой результат обособления части материка и океаническими (вулканические и коралловые острова). Растительный и животный мир первых близок по составу к материковому. На Британских островах и на Сахалине большинство видов сходно с близлежащими районами континента. Это объясняется тем, что острова лишь несколько тысяч лет назад отделились от суши. Однако чем древнее остров и чем более значительна водная преграда, тем больше обнаруживается отличий. На Мадагаскаре нет типичных для Африки крупных копытных:
Сходство зародышевого развития позвоночных
Факт единства происхождения живых организмов был подтвержден на основе эмбриологических исследований. Все многоклеточные животные развиваются из одной оплодотворенной яйцеклетки. В процессе индивидуального развития они проходят стадии дробления, образования двух- и трехслойного зародышей, формирования органов из зародышевых листков.
Сходство зародышевого развития животных свидетельствует о единстве их происхождения.
С особой отчетливостью сходство эмбриональных стадий выступает в пределах отдельных типов и классов. Так, на ранних стадиях развития у зародышей позвоночных (рыбы, ящерицы, кролика, человека) наблюдается поразительное сходство: все они имеют головной, туловищный и хвостовой отделы, зачатки конечностей, по бокам тела — зачатки жабр ( рис. 17 ).
По мере развития зародышей черты различия выступают все более явственно. Причем вначале проявляются признаки класса, к которому относятся зародыши, затем признаки отряда и на еще более поздних стадиях — признаки рода и вида. Эта закономерность в развитии зародышей указывает на их родство, происхождение от одного ствола, который в ходе эволюции распался на множество ветвей.
Гомологичные органы имеют не только сходное строение. Они развиваются из одних и тех же зародышевых листков в процессе индивидуального развития. Такие органы могут выполнять неодинаковые функции, что обусловлено приспособлением к определенным условиям среды обитания. Ласты тюленя гомологичны крыльям птиц, конечностям крота и человека.
Типы деревьев
Не все деревья одинаковы, существуют разные графические изображения, и каждому из них удается включить какую-то особенную характеристику эволюции группы.
Самыми основными деревьями являются кладограммы. Эти графики показывают отношения с точки зрения общих предков (согласно самым последним общим предкам).
Аддитивные деревья содержат дополнительную информацию и представлены в длине ветвей.
Числа, которые связаны с каждой ветвью, соответствуют некоторому атрибуту в последовательности — такой как количество эволюционных изменений, которые испытали организмы. В дополнение к «аддитивным деревьям» они также известны как метрические деревья или филограммы.
Ультраметрические деревья, также называемые дендограммами, являются частным случаем аддитивных деревьев, где вершины дерева равноудалены от корня до дерева.
Эти два последних варианта имеют все данные, которые мы можем найти в кладограмме, и дополнительную информацию. Поэтому они не являются взаимоисключающими, если не дополняют друг друга.
polytomies
Во многих случаях узлы деревьев не полностью разрешены. Визуально говорят, что существует политика, когда новая оставляет более трех ветвей (у более чем двух непосредственных потомков только один предок). Когда у дерева нет политомий, оно считается полностью разрешенным.
Есть два типа политомий. Первыми являются «жесткие» политомии. Они присущи исследовательской группе и указывают на то, что потомки развивались одновременно. Альтернативно, «мягкие» политомии указывают на неразрешенные отношения, вызванные данными per se.
Параллелизмы затрудняют филогенетические реконструкции и часто служат причинами ошибок в них. Неполнота палеонтологической летописи приводит к тому, что в один филогенетический ряд попадают организмы из разных параллельных таксонов. Сходство между ними высокое, а родство отдаленное.
Дикие лошади
Сегодня настоящих диких лошадей практически не осталось. Российский тарпан считается вымершим видом, а лошадь Пржевальского в естественных условиях не встречается. Табуны лошадей, которые пасутся вольно, — это одичавшие одомашненные формы. Такие лошади хоть и быстро возвращаются к дикой жизни, но все же отличаются от истинно диких лошадей.
У них длинные гривы и хвосты, и они разномастные. Исключительно буланые лошади Пржевальского и мышастые тарпаны имеют как бы подстриженные челки, гривы и хвосты.

В Центральной и Северной Америке дикие лошади были полностью истреблены индейцами и появились там только после прибытия европейцев в XV веке. Одичавшие потомки лошадей конкистадоров дали начало многочисленным табунам мустангов, численность которых сегодня контролируется отстрелом.
Кроме мустангов в Северной Америке есть два вида диких островных пони – на островах Ассатиг и Сейбл. Полудикие табуны лошадей камаргу встречаются на юге Франции. В горах и болотах Британии тоже можно встретить некоторых диких пони.

Доказательства эволюции живой природы
Эволюционная теория со времени Ч. Дарвина обогатилась данными палеонтологии, анатомии, биогеографии и других наук.
Палеонтологические доказательства
Ископаемые вымершие организмы находили и до создания теории эволюции. Собраны окаменевшие отпечатки, пропитанные кремнеземом, известняком остатки, следы жизнедеятельности. К редким находкам относятся хорошо сохранившиеся организмы, вмерзшие в лед, «законсервированные» в вечной мерзлоте, заключенные в янтарь.
Найдены промежуточные формы (переходные), сочетающие признаки эволюционно древних и более молодых групп организмов. К таковым относятся отпечатки растений псилофитов, окаменевшие остатки стегоцефалов, археоптерикса, зверозубых ящеров (Таблицы 1, 2 и 3, рис. 2), «живое ископаемое» — рыба латимерия (рис. 1).
- Тело — слоевище, не дифференцированное на органы;
- Ткани отсутствуют.
- Стебель с чешуйками;
- Устьица в кожице стебля и листьев;
- Корневище с ризоидами;
- Есть покровные, механические и проводящие ткани.
Рис. 1 Кистеперая рыба латимерия
Таблица 2.
Археоптерикс
- Длинный хвост с позвонками;
- Пальцы с когтями;
- Наличие зубов;
- Отсутствие киля;
- Брюшные ребра.
- Внешнее сходство;
- Наличие крыльев;
- Оперение;
- Сходство задних конечностей с цевкой.
Зверозубый ящер
- Внешний облик;
- Расположение конечностей по бокам туловища, а не под ним.
- Строение черепа, позвоночника и конечностей;
- Деление зубного ряда на клыки, резцы и коренные зубы.
Значительным доказательством эволюции считается то, что в более ранних слоях встречаются просто устроенные организмы, по сравнению с находками в более поздних субстратах. Останки видов, обитавших в одни геологические эпохи, позже исчезают и больше не появляются. Это означает, что виды возникают в ходе эволюции и вымирают в последующие эры и периоды.
Сравнительно-анатомические доказательства
Находки ископаемых останков организмов часто имеют общее происхождение и сходство с современными. Изучение строение ныне существующих и исчезнувших позвоночных животных выявило общие черты строения:
- Двусторонняя симметрия.
- Твердый скелет.
- Полости тела.
- Сходное строение нервной системы.
Сравнение ископаемых останков предков лошади с анатомическим строением современных животных позволило основателю эволюционной палеонтологии В. О. Ковалевскому доказать поступательный характер эволюционного процесса. Ученый создал филогенетический ряд, показывающий эволюцию предков лошади (рис. 3).
Рис. 3. Филогенетический ряд, предложенный В. О. Ковалевским
К сравнительно-анатомическим доказательствам относят строение цветка покрытосеменных растений. Изучение этого органа у представителей разных семейств, родов и видов дает убедительные свидетельства сходства. История развития и действие факторов среды накладывают отпечаток на строение органа. Однако у покрытосеменных, при удивительном разнообразии формы и окраски, цветки имеют чашечку, лепестки венчика, пестики и тычинки.
Конечности у наземных позвоночных животных имеет общий план строения, хотя могут быть не похожи. Изучение анатомического строения доказывает, что это разные варианты пятипалой конечности (рис. 4).


